
Cell-Penetrating Peptides (CPPs)
The delivery of therapeutic agents into the intracellular environment to interact with intracellular macromolecules is a prerequisite for the therapeutic application of many biological drugs. However, the plasma membrane of cells acts as a biological barrier that prevents most hydrophilic macromolecules from entering the cell, which severely limits the development and application of these molecules in disease treatment. Therefore, how to efficiently deliver these drugs into cells has always been a major challenge in drug development. Currently, the primary methods used to guide macromolecules across biological membranes include **Cell-Penetrating Peptides (CPPs)**, liposomes, virus-like particles, and nanoparticles. Among these, CPPs act as vectors by binding to their cargo, typically entering cells via receptor-independent mechanisms. They are generally less than 30 amino acids in length, rich in basic amino acids, and possess an overall positive charge.
The first human cell-penetrating peptide discovered was the **TAT transactivation factor** derived from Human Immunodeficiency Virus Type 1 (HIV-1 TAT). It penetrates biological membranes in a non-destructive and highly efficient manner to enter cells.
A key feature of cell-penetrating peptides (CPPs) is their capacity to deliver various cell-impermeable biologically active macromolecules into cells, including small molecule compounds, oligonucleotides, peptides, peptide nucleic acids (PNA), proteins, plasmid DNA, siRNA, and 200 nm liposomes or iron oxide nanoparticles. This unique property offers immense potential for the delivery of intracellularly targeted therapeutics.
The outstanding advantages of CPPs as delivery vectors lie in their low cytotoxicity and the predictability of cargo release across various cell lines. Although CPPs can enter cells through multiple pathways depending on the cell type, their practical applications are most extensively documented in the delivery of oligonucleotides (ONs), proteins, and plasmid DNA in vitro and in vivo.
Translocation Mechanisms of CPPs
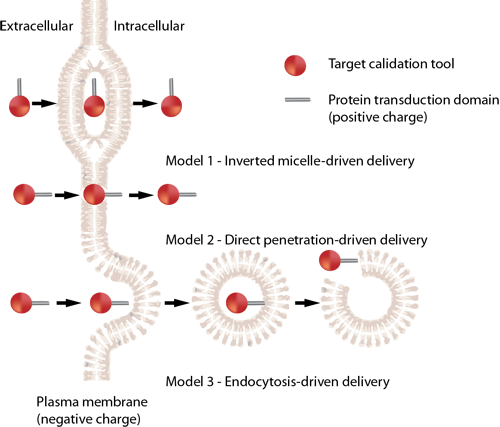
Different cell-penetrating peptides exhibit distinct internalization mechanisms. The specific pathway utilized by a given CPP depends on several parameters, including molecular size (e.g., cargo properties), temperature, cell phenotype, and the stability of the peptide both inside and outside the cell environment. The exact internalization mechanisms of cell-penetrating peptides remain a subject of ongoing discussion. Currently, the most widely accepted pathways can be summarized into the following three basic models:
■ Table 5: Representative Sequences and Origins of Well-Characterized CPPs
| Name | Origin | Sequence |
|---|---|---|
| Tat family | ||
| Tat (48-60) | HIV-1 protein | GRKKRRQRRRPPQ |
| Oligoarginine | Tat derivative | Rn |
| Penetratin family | ||
| p-Antp | Antennapedia homeodomain | RQIKIWFQNRRMKWKK |
| pIsI | Igl-1 homeodomain | RVIRVWFQNKRCKDKK |
| Chimeric CPPs | ||
| Transportan | Galanin-mastoparan | GWTLNSAGYLLGKINLKALAALAKKIL |
| MPG peptides | ||
| P-beta | gp41-SV40 | GALFLGFLGAAGSTMGAWSQPKKKRKV |
| P-alpha | gp41-SV40 | GALFLAFLAAALSLMGLWSQPKKKRKV |
| Pep-1 | Trp-rich motif-SV40 | KETWWETWWTEWSQPKKKRKV |
Case Study: Molecular Mechanism of HIV TAT Translocation
Cell-penetrating peptides, particularly HIV TAT, can enter cells via both direct translocation and endocytosis. The HIV TAT protein transduction domain can be engineered into an efficient drug delivery vector. However, the exact molecular mechanisms underlying how cell-penetrating peptides cross biological membranes remain a subject of active scientific exploration.
To uncover how HIV TAT transitions between direct membrane penetration and endocytosis, a research team led by Gerard Wong investigated how HIV TAT interacts with biological membranes, actin filaments, and specific proteoglycan receptors under varying experimental conditions. Their findings provided multiple lines of structural evidence clarifying this complex biophysical phenomenon.
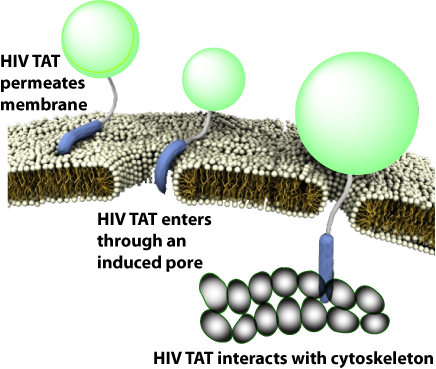
Intriguingly, TAT can undergo entirely distinct biophysical reactions with the same sequence under varying cellular environments. Through interactions with cell membranes, actin filaments, and proteoglycans, it induces diverse localized topological curvatures and drives distinct transport processes.
The core mechanism governing the cell membrane translocation of CPPs is intrinsically tied to their structural amino acid distribution. If a hydrophobic spacer group is incorporated into a hydrophilic CPP sequence, the structural mechanism of transmembrane delivery can be significantly shifted. For instance, a simple CPP sequence like polyarginine (polyR) can induce an inward-directed curvature on the biological membrane, forcing extracellular components to enter the internal cavity, thereby forming an endocytic vesicle. Concurrently, the peptide can induce both positive curvature and negative curvature on the lipid layer. When the curvature indices balance in opposite orthogonal directions, a stable saddle-splay curvature is established, which is thermodynamically favorable for creating a transient transmembrane pore, allowing direct penetration of small molecules.
If the sequence structural hydrophobicity favors the generation of negative Gaussian curvature, why is the translocation capacity of HIV TAT closely linked to its specific arginine content or structural layout? The underlying reason is that CPPs efficiently leverage the water-insoluble guanidinium headgroups of arginine to generate the necessary **saddle-splay curvature**. Simple linear permutations of amino sequences may only generate localized geometric anomalies on the membrane surface, preventing the spatial coordination required for complete macro-pore opening. Because different CPP types feature distinct sequence arrangements, TAT can efficiently initiate transmembrane delivery under specific conditions even without the support of formal cell-surface receptor complexes.